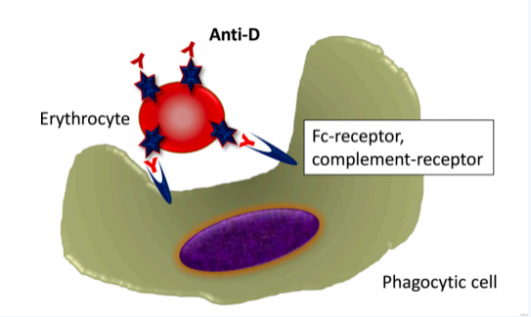
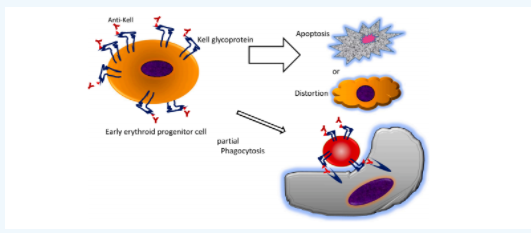
虽然这三种非经典的新生儿溶血的模式并不常见,但是对于我们理解IgG抗M、抗Ge、抗Kell的溶血,无法用经典HDN解释其临床表型和实验室特性,是十分有益的。特别是第三种克隆逃逸,之前已有学者发现,在肺癌环境下HLA特异性等位基因会丢失(McGranahan et al.,2017,Cell 171,1259-71),而由抗CD38诱导的红细胞膜上CD38抗原的丢失(Harold C. Sullivan et al.,2017 Blood 129 (22): 3033–37.)。
本稿件由协会免疫血液学专业委员会提供
作者:朱自严
原文:点击此处可下载查看(Ohto Hitoshi .et,al. Three non-classical mechanisms for anemic disease of the fetus and newborn, based on maternal anti-Kell, anti-Ge3, anti-M, and anti-Jr a cases. Transfus Apher Sci 2020 Oct;59(5):102949. doi: 10.1016/j.transci.2020.102949.)